- 細菌の形質転換
染色体上に存在する遺伝子の化学的本体がDNAであることを明らかにした最初の実験は1928年Griffithが肺炎双球菌(Pneumococus)を使った実験で、遺伝学の歴史の上からも、画期的な業績である。この実験で、遺伝的形質を規定する物質が、細菌細胞の間で伝達されることが初めて明確に示され、細菌の遺伝学の基礎を築いたばかりでなく、その後の分子遺伝学の発展の上でも重要な発見となった。
肺炎双球菌には菌体表面に莢膜をもち、滑らかなコロニーを形成し、また強い病原性をもつS型と、莢膜を失ってでこぼこのあるコロニーを作り、また病原性を失ったR型のものがある。病原性のあるS型をマウスに注射すると、マウスは肺炎で死ぬが、病原性のないR型の菌を注射してもマウスは死なない。S型の菌を熱処理して殺すと、これを注射してもマウスは死なずに生存する。
しかし、熱処理をして殺したS型菌と、生きたR型菌を混合してマウスに注射すると、マウスは肺炎にかかって死んだ。このことは熱処理されたS型菌の遺伝物質がR型菌に伝達されて、R型菌がS型菌に変化したことを示すもので、この現象を形質転換(transformation)という。この遺伝物質がDNAであることは1944年にAvery、MacLeodおよびMcCartyによって明らかにされた。「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- 細菌の性と染色体組換え
高等動植物と異なり、細菌やウイルスは肉眼では観察することができない。したがって、その形態や色彩など目に見える形質を対象とした遺伝学的研究を、これらの微生物にそのまま適用することはできない。大腸菌、枯草菌、サルモネラ菌などの細菌は寒天培地の上で分裂、増殖してコロニーを形成する。これらの細菌の分裂増殖のためには、培地成分として種々の栄養物質を必要とし、またこれらの栄養物質を利用して、自分で多くの酵素を生成することも必要である。
細菌の遺伝学は、このように細菌の増殖に必要な栄養物質(たとえばアミノ酸やビタミン類)の要求性や、ストレプトマイシン、ペニシリンなど抗生物質に対する抵抗性の性質、さらには細菌に寄生するバクテリオファージに抵抗性の性質などを指標として、これらが遺伝的に伝わることが見いだされ、その遺伝様式を調べることから始まった。
多くの細菌はグルコースと無機塩のみの最小培地でも増殖できる。しかし、この細菌にニトロソグアニジンなどの突然変異誘発物質(変異原)を作用させると、この最小培地では成育できない突然変異が生じる。これらの突然変異の多くは、この最小培地に、あるアミノ酸やビタミンを添加すると増殖するようになり、このようにして種々の栄養要求性突然変異体を分離することができる。抗生物質やファージに抵抗性の突然変異体も同じようにして分離することができる。
このようにして多くの突然変異体が分離され、これらを利用した遺伝学の研究に発展したのは、すでに述べた細菌にも雌雄の性があり、雄と雌の細菌の接合が起ることが明らかになったためである。大腸菌には雄と雌の細菌があり、雄の細菌は細胞質のF因子(性決定因子)をもっている。F因子が染色体に付着すると、雌の細菌に雄の染色体を移す能力をもつ。雄(F+)と雌(F-)の細菌が接合して、細い橋によってつながると、雄の環状の染色体が1か所から切断されて、F因子が挿入された箇所から染色体の複製が始まる。こうして新しくできた染色体は、その一端から細い橋を通って雌の細胞の中に移動する。37℃では染色体全体が移動するのに約90分を必要とする。
しかし、途中で接合が破れると、雄の染色体の一部だけしか雌の細胞に入らず、雌の細胞の中で入ってきた雄の染色体の一部と雌の染色体との間の組換えが起る。このようにして、雄と雌の細菌の接合を利用して、雄の染色体上の遺伝子の移動する頻度をもとに、大腸菌の染色体地図が作成された。F因子のように細胞質内に存在し、遊離の状態と染色体図に組込まれた状態の二つの状態をとることのできる因子をエピソーム(episome)と呼び、エピソームにはF因子のほか、薬剤抵抗性を支配するR因子(resistant factor)などもある。
このようにF+菌とF-菌が接合し、F因子によって組換え体ができる頻度はF+菌あたり10-4くらいと非常に低いが、これよりも1000倍も高率に組換え体をつくる突然変異体としてHfr(high frequency of recombination)が発見された。このHfr菌ではF因子が最初からF+菌の染色体上の一定の位置に付着しているものと考えられている。「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- 細菌の形質導入
大腸菌には、大腸菌の細胞の中で増殖し、細胞を破壊して、さらに他の大腸菌に感染して増殖を続けるウイルスがある。一般に細菌に感染して、その中で増殖するウイルスをバクテリオファージ(bacteriophage)と呼ぶ。バクテリオファージには細菌に感染して、その中で増殖して細菌を殺し、次々と他の細菌に感染を続ける毒性ファージ(virulent phage)と、感染した細菌内で、細菌の染色体に組込まれ(溶原化という)、細菌と同じ速度で分裂するが紫外線などの照射を行うと、ファージは染色体から離脱して、毒性ファージのように細菌内で増殖して、細胞を破壊して外部に出て、他の細菌に感染する溶原性ファージ(lysogenic phage または temperate phage)とがある(図1・110)
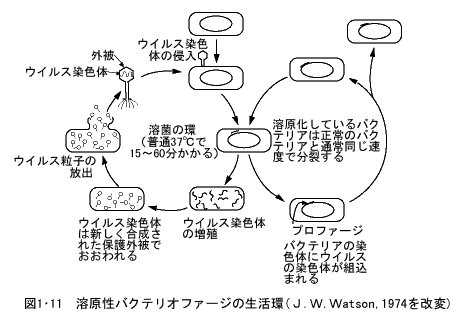
毒性ファージとしては、T2、T4、T6などTシリーズのファージがよく知られており、溶原性ファージとしてはλファージがもっともよく知られている。ファージの頭部には染色体DNAが存在し、尾部で細菌の表面に接着すると、尾部から酵素が出て細菌の外膜をこわし、ファージ頭部のDNAが尾部を通って細菌内に注入され、タンパク質の外被は細菌外に取り残される。
バクテリオファージが遺伝学的に有用な材料として取り扱われたのは、λファージなどの溶原性ファージで、宿主細菌の染色体に組込まれて、あたかも細菌の染色体の一部として増殖し、それが再び染色体から離脱して単独で行動することができる。そのため、λファージに別の遺伝子を組込ませて感染させると、その遺伝子を細菌にもち込むことができる遺伝子の運び屋(ベクター)としての役割があるからである。
さらにまた、T4ファージのような毒性ファージには、寒天上に成育した大腸菌に感染させると、大腸菌の細胞層の中にファージが次々に感染して大腸菌を殺し、大腸菌のない透明な室間(プラークという)を作る。そしてこのプラークの形に、大きさや透明度の異なる突然変異のファージがある。また、感染する大腸菌にもT4ファージに抵抗性の大腸菌があり、この抵抗性の大腸菌に感染するT4ファージの突然変異体(宿主域突然変異体)が存在する。これらの異なった性質をもつ突然変異のファージを、同じ大腸菌に重複感染させると、大腸菌内でそれぞれのファージの染色体は複製を繰り返し行い、できた染色体間に組換えを起こす。このようにしてできた組換え染色体をもつファージは、新しい遺伝子の組合せをもつファージとなる。バクテリオファージではこのような2種の異なったファージを同時感染させて、その染色体の組換え頻度をもとにファージの染色体地図が作成されている。
大腸菌のような細菌の染色体の一部がファージによって一つの細菌細胞から他の細菌細胞に運びこまれ、これによって、運び込まれた遺伝子による新しい形質がその細胞に生じる現象を形質導入(transduction)という。導入される遺伝形質としては栄養要求性や薬剤抵抗性のほか抗原性、糖分解能、運動性など種々の形質がある。また、大腸菌のP22やP1というような多くのバクテリオファージはどのような遺伝形質でも導入することができ、これを普遍導入(generalized transduction)という。これに対して、同じ大腸菌に感染するファージでも、λファージのようにあるきまった遺伝子しか導入しないものを特殊導入(specialized transduction)という。
細菌やファージを用いた微生物の遺伝学は、細胞が単細胞で単純であり、集団として取扱い易く、統計的処理や定量的処理が容易であり、また世代時間がきわめて短いなど遺伝学的研究の材料として非常に有利な性質をそなえている。このため、遺伝子の本体としてのDNAを中心としたその後の分子遺伝学の発展に主導的役割を果した。「基礎遺伝学」(黒田行昭著;近代遺伝学の流れ)裳華房(1995)より転載
- 遺伝子の微細構造
細菌やファージの遺伝学が進み、大腸菌を宿主とするT4ファージを用いて、遺伝子の微細構造を明らかにする実験が行われた。これはBenzer(1955)が、T4ファージとその突然変異体であるγIIという系統を用いて行ったものである。正常の T4ファージは大腸菌のK株細胞の上にまくと、細菌の細胞を溶かしてプラーク(溶菌班)を形成する。ところがγIIファージは大腸菌K株の上ではプラークを形成することができない。このγIIの突然変異を8種類とって、その2種類ずつを一緒にして大腸菌K株にまくと、時にはプラークを形成するものもある。これは2種類のγII系統がγII遺伝子の中の異なった領域に突然変異を起しており、混合感染させた大腸菌の菌体内で染色体の組換えを起し、遺伝子の欠失した領域を互いに補い合って正常のファージを生じ、これがプラークを作ったものと考えられた。
このことからBenzerは一つの遺伝子と考えられるものも、さらにいくつかの小単位に分けられることがわかり、つぎのような単位を提唱した。- レコン(recon):染色体の乗換えを起すことのできる最小単位で、その距離は0.01~0.02%くらいである
- ミュートン(muton):突然変異を起すことのできる最小単位で、レコンより少し大きく、0.05%くらい。
- シストロン(cistron):レコンがいくつか集まってできた単位で、1種の生理的単位。一般に遺伝子と考えられるのはこのシストロンに相当する。
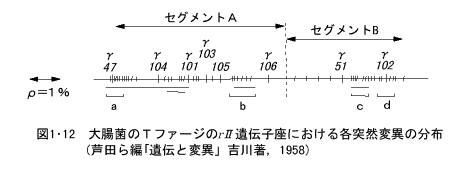
T4ファージのγII遺伝子座における各突然変異の分布を示すと図1・12のようになる。
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- DNAの構造と複製
遺伝子の本体がDNA(一部RNA)であることは、肺炎双球菌で病原性の強い菌の遺伝物質が熱処理後も他の病原性を失った菌に移って、強い病原性を示し、この遺伝物質がDNAであることからも明らかである。さらに、ファージに感染した細菌の細胞内に入るのはファージの核酸だけで、被膜のタンパク質は菌体外に残され、菌体内に入った核酸は複製して完全な多数のファージとなって放出され、別の細菌に感染することによっても証明される。
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- DNAの構造
DNAの化学的構造を明確にしたのは、WatsonとCrick(1953)である。DNA分子は二重らせん構造がその基本的な骨格で(図1・13)、デオキシリボースという五炭糖の5'Cと3'Cがリン酸と交互に連なった鎖が2本あり、この鎖が相対して平行に並んで走り、その五炭糖にはアデニン(A)、グアニン(G)、シトシン(C)、チミン(T)という4種の塩基が、一方の鎖のAに対しては他の鎖のT、Gに対してはC というように結合して2本の鎖をはしごの段のようにくっつけた構造をしている(図1・13)。
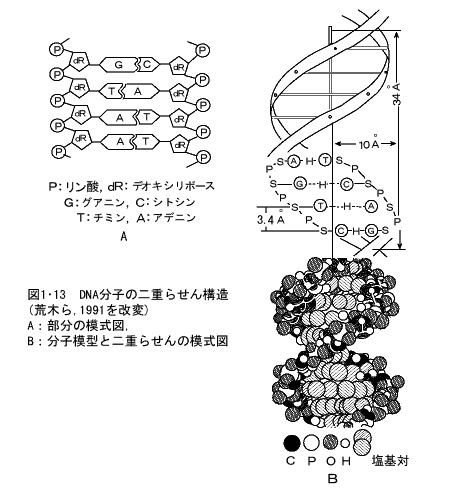
このAとT、GとCの塩基は相補的な関係にあって、2本の鎖の塩基の配列を規定しており、この4種類の塩基の配列順序が遺伝情報となっている。このDNA分子を塩基、リン酸、五炭糖に分解して考えると、一つの塩基に五炭糖の1'Cが結合したものをヌクレオシド(nucleoside)といい、この五炭糖の5'Cにリン酸1分子が結合したものをヌクレオチド(nucleotide)という。したがってDNAの各1本の鎖は塩基の種類を異にした4種のヌクレオチドが連なったものといえる。2本鎖の相対する塩基は上に述べたように、相補的な関係にあるため、種々の生物のDNAを構成する塩基組成の構成比はA:TおよびG:Cの比はいずれも1であり、A+T/G+Cの比に、それぞれの生物のDNAの特徴がみられる。
このDNAの2本の鎖がらせん状にねじれており、4種の塩基を内部に包み込んだ構造になっている。このDNAの二重らせん構造が重要な意味をもつのは、つぎのようにDNAの主な機能をよく説明することができるからである。- DNA分子は自分と同じ塩基配列をもった分子を、正確に忠実に複製できる。
- DNA分子の塩基配列によって形成される遺伝情報は、伝令RNA(メッセンジャーRNA、mRNA)によって正確に写しとられ(転写)、その塩基配列が3個ずつトリプレットとなって一つのアミノ酸を指定して、きまったアミノ酸の配列をもつタンパク質を合成する(翻訳)。
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- DNAの複製
細胞が分裂、増殖するとき、細胞の核内では染色体に組込まれたDNAが自分と同じ塩基配列をもつ二つの分子に複製される。これは細胞サイクルのDNA合成期(S期)に行われる。この際、染色体に何重ものらせん状になって組込まれていたDNA(1・5・3染色体の微細構造参照)の二重らせんが一定の場所 (複製開始点) からほどけて、一本ずつの鎖に開裂する(図1・14)。
そして、それぞれ一本ずつの鎖のA、T、G、Cの塩基に対応して、AにはT、TにはA、GにはC、CにはGのヌクレオチド(塩基+五炭糖+リン酸)を選択的に結合して、隣接したヌクレオチドの五炭糖の3'Cとリン酸とが結合して、ヌクレオチドの長い鎖ができる(図1・14C)。この新しくできたDNA鎖はもとのDNA鎖の塩基配列と相補的な塩基配列になっており、もとのDNAの2本鎖がそれぞれ鋳型となって、それに相補的な塩基配列をもった新しい鎖をつくるので、もとのDNA1分子から塩基配列のまったく同じ2分子のDNAができる(図1・14D)。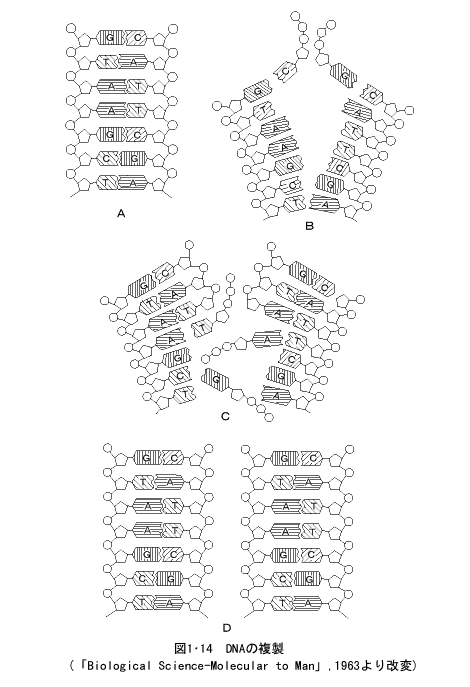
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- DNAからのタンパク質合成
DNA分子の4種類の塩基の配列順序が遺伝情報といわれるもので、二重鎖の中の一本(これをマスター鎖という)が鋳型となって、その塩基の配列に対応した塩基配列(Aに対してウラシル[U]、Tに対してA、Cに対してG、Gに対してC)をもった伝令RNA (メッセンジャーRNA、messenger RNA、mRNA) が合成される。これをDNAの遺伝情報の転写(transcription)という (図1・15)。
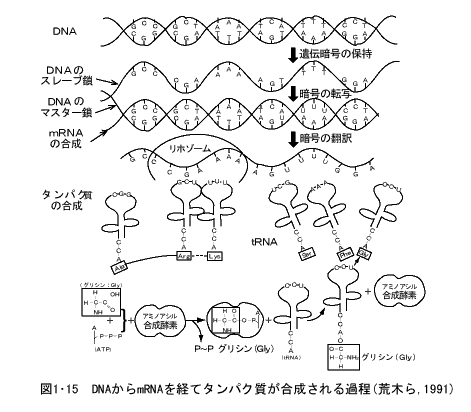
RNAの分子はDNAと同じように4種類の塩基と五炭糖、リン酸とからなる。しかし、DNAと異なるのは、塩基の中のT(チミン)の代わりにU(ウラシル)が使用され、また五炭糖はDNAのデオキシリボースの代りにリボースが使われていることとDNAは二重鎖であったが、RNAは一本鎖であることである。
DNAからmRNAが転写されるとき、RNA合成酵素(RNAポリメラーゼ)IIが、鋳型になるDNAの特異的な塩基配列(約50塩基対)を読みとってそこに結合し、転写を開始し、しだいにDNAの遺伝情報を読み取ってRNA鎖を伸長させる。ここまでの過程は、細胞の核の中で起る現象である。
このようにしてできたmRNAは核膜の孔を通って細胞質に移動し、細胞質中のリボソーム(ribosome)に結合し、これを足場として、一方の端から、三つずつの塩基配列(これをトリプレット[triplet]、またはコドン[codon]という)に対応した相補的な三つの塩基(これをアンチコドン[anticodon]という)をもつ転移RNA(トランスファーRNA、[transferRNA、tRNA])が、それぞれ固有のアミノ酸を運んでくる。
このtRNAはクローバの葉の形をした約80個のヌクレオチドからなる分子(分子量約25,000)で一方の端にはmRNAのコドン対するアンチコドンの三つの塩基があり、他方の端にはそれぞれのアンチコドンに特有なアミノ酸をアミノアシル合成酵素の働きにより結合している。
mRNAの三つずつの塩基を読みとって運ばれてきたtRNAはmRNAと結合し、他端のアミノ酸が隣接のtRNAのアミノ酸と結合して、tRNAより離れ、アミノ酸のみのペプチドを作る。これはしだいに伸長してタンパク質となる。このmRNAの遺伝情報にもとづいて、特有なアミノ酸配列をもつタンパク質が合成される過程を翻訳(translation)という。
微生物からヒトにいたるあらゆる生物で起るDNAからタンパク質合成の道筋はほとんど上記のような基本的な原理にもとづいて行われ、DNA中に塩基配列としてたくわえられた遺伝情報が転写の過程を経てmRNAに移され、さらに翻訳の過程を経て正確に一定のアミノ酸順序をもつタンパク質の合成が行われることになる。このDNAからタンパク質合成までの過程をセントラルドグマ(central dogma)と呼ぶ。「基礎遺伝学」(黒田行昭著;近代遺伝学の流れ)裳華房(1995)より転載
- スプライシング
細菌などの微生物では、mRNAの遺伝情報がそのままタンパク質のアミノ酸配列として翻訳されるが、高等生物などの真核生物(eukaryote)の細胞では、mRNAの遺伝情報がそっくりそのままリボソーム上でタンパク質のアミノ酸配列として翻訳されることはむしろ稀で、mRMAはいくつかの部分に切断され、そのある部分(イントロン、intron)は除去されて、残りの部分(エクソン、exon)のみが再結合して、新しいmRNAが形成される。この過程をスプライシング(splicing)という(図1・16)。
このイントロンが捨てられ、エクソンが再結合する場合に、エクソン断片の再結合に異なったしかたがあり、もとの一つのmRNA前駆体から異なった組合せのエクソンの再結合により、複数の異なったmRNAが形成されることがある。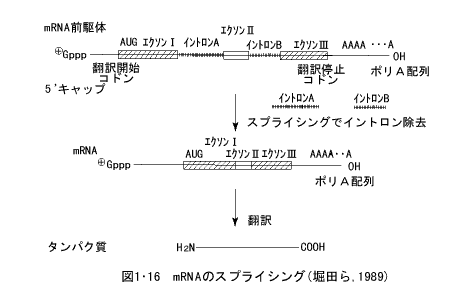
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- 遺伝暗号
mRNAの三つずつの塩基(コドン)が、どのようなアミノ酸をもつtRNAに対応しているかを示したものが遺伝暗号(genetic code)と呼ばれるものである。遺伝暗号は図1・17に示したようにmRNAの第1、第2、第3番目の三つの塩基に対応するそれぞれのアミノ酸が示されている。この表によって明らかなように、同一のアミノ酸に対していくつかの異なったコドンが対応している。とくに1番目の塩基と2番目の塩基が同じ場合には、3番目の塩基が異なっても同じアミノ酸に対応することが多く“ゆらぎ”(wobble)という現象がみられる。3番目の塩基がUとC、またはAとGというようにピリミジン塩基またはプリン塩基どうしの置換は、対応するアミノ酸が同じものが多い。このようにいくつかの異なったコドンが一つのアミノ酸に対応する現象を遺伝暗号の縮重(デジェネラシー、degeneracy)といわれる。
遺伝暗号の中にはAUGというタンパク質合成の開始コドン(initiation codon)があり、タンパク質合成が開始されるときには、ここにホルミルメチオニンと結合したtRNAが結合する。このホルミルメチオニンはタンパク質合成が終了したとき除かれる。またUAA、UAG、UGAというタンパク質合成の終止コドン(termination codon)があり、これには対応するtRNAがないためにタンパク質合成はこの前で終了する。2文字目 U C A G 3文字目 1文字目 U UUU フェニルアラニン UCU セリン UAU チロシン UGU システイン U UUC フェニルアラニン UCC セリン UAC チロシン UGC システイン C UUA ロイシン UCA セリン UAA 終止コドン UGA 終止コドン A UUG ロイシン UCG セリン UAG 終止コドン UGG トリプトファン G C CUU ロイシン CCU プロリン CAU ヒスチジン CGU アルギニン U CUC ロイシン CCC プロリン CAC ヒスチジン CGC アルギニン C CUA ロイシン CCA プロリン CAA グルタミン CGA アルギニン A CUG ロイシン CCG プロリン CAG グルタミン CGG アルギニン G A AUU イソロイシン ACU スレオニン AAU アスパラギン AGU セリン U AUC イソロイシン ACC スレオニン AAC アスパラギン AGC セリン C AUA イソロイシン ACA スレオニン AAA リジン AGA アルギニン A AUG メチオニン(開始コドン) ACG スレオニン AAG リジン AGG アルギニン G G GUU バリン GCU アラニン GAU アスパラギン酸 GGU グリシン U GUC バリン GCC アラニン GAC アスパラギン酸 GGC グリシン C GUA バリン GCA アラニン GAA グルタミン酸 GGA グリシン A GUG バリン GCG アラニン GAG グルタミン酸 GGG グリシン G 「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
