- 染色体の数と形
“The history of the earth is recorded in the layers of its crust; The history of all organisms is inscribed in the chromosomes”(地球の歴史は地殻の層にあり、すべての生物の歴史は染色体にきざまれている)木原 均(1946)。
これはコムギの遺伝学的研究では、世界的に有名な木原 均博士の残された言葉である。約30億年前地球上に生物が誕生し、原始生物から高等な動物植物に進化した長い歴史は、生物体を構成している細胞の核の中にある染色体にきざみ込まれている。生物は個体が死滅しても、細胞の中の染色体(遺伝子)は、生殖細胞を通じて子孫に伝わり、染色体の中に組込まれた遺伝子が生物体のすべてのタンパク質や酵素、その他の成分を作り、細胞としての特色ある機能を営ませる。
1865年にメンデルの法則が発見され、生物のもっている形質がどのように子孫に伝わるかが科学的な実験事実に基づいて発表されたわけである。しかし、このような生物の表現型の遺伝を支配する遺伝子そのものが、細胞核の中にある染色体(chromosome)という構造物の中に存在することが明らかになったのは、19世紀の末にWaldeyer(1888)が染色体の命名を行った後である。これは、光学顕微鏡の発達、改良と深い関係にあり、細胞の内部構造の観察から核や細胞質、さらには細胞が分裂するときに姿を現す染色体の発見につながったのである。
染色体の数と形は、それぞれの生物種によって決まっている。少ないものではウマノカイチュウやアオカビの2本から、多いものではヤドカリの254本に達するものもある。体を作っている細胞(体細胞 : somatic cell)では、同じ大きさで同じ形をした染色体(相同染色体)が2本ずつペアで存在し、このペアが生物種によって決まった数だけ存在している。したがって1本ずつ異なった染色体のセット(これをゲノムという)が、2セット存在しているともいえる。この2セットの染色体をもつ細胞を二倍体細胞(diploid)といい、生殖細胞(germ cell)ができるときには、この2セットの染色体の組が、減数分裂(meiosis ; meiotic division)により、それぞれ1セットずつ生殖細胞に入るので、生殖細胞は1ゲノムの染色体をもち半数体(haploid)という。生殖細胞が受精すると再び2セットの染色体に戻る。
染色体の中の1ペアの染色体は性染色体(sex chromosome) と呼ばれ、多くの生物では性染色体の組合せが雄と雌によって異なり、性の決定にあずかっている。残りのすべての染色体は常染色体(autosome)と呼ばれ、雄と雌とでは差違はない。
表1・2に主な動物と植物について、その染色体の数と性染色体の構成を示す。常染色体の数は同じ大きさと形をしたものが2本ずつあり、性染色体は雌がX染色体2本でXX、雄がX染色体とY染色体のヘテロの組合せになっているものと、逆に雄がZ染色体2本でZZ、雌がZ染色体とW染色体のヘテロの組合せになっているものがある。雌ヘテロ型の生物はニワトリやハトなどの鳥類やカイコ、チョウなどの鱗翅目の昆虫に見られるが、他の大部分の生物では雄ヘテロ型になっている。表1・2:主な動植物の染色体数と性染色体(荒木ら「現代生物学図説」、1991)
A.動物 B.植物 種名 常染色体+性染色体 種名 常染色体+性染色体 ゾウリムシ Paramecium aurelia 2n:30~40 アオカビ Penicillium sp. n:2 ウマノカイチュウ Ascaris megalocephala 2n:2 アカパンカビ Neurospora crassa n:7 ミツバチ Apis mellifera 2n:16 スフェロカルプス Sphaerocarpus donnellii(コケ類) n:♀7+X,♂7+Y カイコガ Bombyx mori 2n:♀54+ZW,♂54+ZZ ストロブマツ Pinus strobus 2n:24 バッタ Melanoplus differentialis 2n:♀22+XX,♂22+XO イチョウ Ginkgo biloba 2n:24 ショウジョウバエ Drosophila melanogaster 2n:♀6+XX,♂6+XY キャベツ Brassica oleracen 2n:18 ザリガニ Cambarus clarkii 2n:200(?) スイカ Citrullus vulgaris 2n:22 ウナギ Anguilla anguilla 2n:36(?) カボチャ Cucumis sasivus 2n:14 アマガエル Hyla sp. 2n:24(?) コーヒーノキ Coffea arabica 2n:44 ニワトリ Callus domestica 2n:♀76+ZW,♂76+ZZ アサ Cannabis sativa ♀18+XX,♂18+XY ハト Columba livia 2n:♀78+ZO,♂78+ZZ カナムグラ Humulus japonicus ♀14+XX,♂14+Y1XY2 ウシ Bos taurus 2n:♀58+XX,♂58+XY タカイチゴ Fragaria elatior ♀40+ZW,♂40+ZZ ウマ Equus caballus 2n:♀62+XX,♂62+XY サンショウ Xanthoxylum piperitum 2n:♀68+XX,♂68+XO ブタ Sus scrofa 2n:♀38+XX,♂38+XY オニドコロ Dioscorea gracillima 2n:♀18+XX,♂18+XY ネコ Felis calus 2n:♀36+XX,♂36+XY タバコ Nicotiana tobacum 2n:48 イヌ Canis familiaris 2n:♀76+XX,♂76+XY バナナ Musa paradisiaca 22,33,44,55,77,88 ハツカネズミ Mus musculus 2n:♀38+XX,♂38+XY カラスムギ Avena sativa 2n:42 シロネズミ Rattus norvegicus 2n:♀40+XX,♂40+XY パンコムギ Triticum vulgare 2n:42 テンジクネズミ Cavia porcellus 2n:♀62+XX,♂62+XY イネ Oryza sasiva 2n:24 カイウサギ Oryctolagus cuniculus 2n:♀42+XX,♂42+XY トウモロコシ Zea mays 2n:20 アカゲザル Macaca mulatta 2n:♀40+XX,♂40+XY タカサゴユリ Lilium formosanum 2n:24 ゴリラ Gorilla golirra 2n:♀46+XX,♂46+XY ソラマメ Vicia faba 2n:12 チンパンジー Pan troglodytus 2n:♀46+XX,♂46+XY タマネギ Allium cepa 2n:16 ヒト Homo sapiens 2n:♀44+XX,♂44+XY Haplopappus gracilis(キク科) 2n:4 ヒトの場合は染色体数は46本で、この中で常染色体が22対で44本、性染色体は女性がXX、男性がXYとなっている。ヒトの常染色体を長さの大きいものから順に並べると、動原体(紡錘糸の付着する場所)が中央にあるもの(中部動原体型:metacentric)が3対(A群)、やや端に片寄ったもの(次端部動原体型:acrocentricまたはsubtelocentric)が2対(B群)、中部に近いもの(次中部動原体型:submetacentric)が7対(C群)、端に片寄ったもの(端部動原体型:elocentric)が3対(D群)、中部に近いものが3対(E群)、中央にあるものが2対(F群)、端に片寄ったものが2対(G群)あり、全部で22対となっている。
普通の光学顕微鏡では、ほぼ同じ大きさで同じ形の染色体を個々に識別することは困難なため、キナクリンマスタード(Qバンド法)やギムザ(Gバンド法)などの染色体分染法を用いると、染色体に染まる部分と、染まらない部分の縞模様を作り出す。これがそれぞれの染色体に特有な縞模様を示すため、個々の染色体を識別することができる。「基礎遺伝学」(黒田行昭著;近代遺伝学の流れ)裳華房(1995)より転載
- 生殖細胞の染色体
生殖細胞は生物の発生のある決まった時期に、体の細胞の一部が普通の細胞の分裂とは異なった分裂をして生殖細胞が形成される。この細胞分裂は減数分裂(meiosis)と呼ばれ、連続2回の分裂があり、これによって細胞の中にある染色体の数が半分になる。この半分になるなり方は、上に述べたように体細胞では2本ずつペアで存在した同じ大きさで同じ長さの相同染色体が1回目(または2回目)の分裂の際にそれぞれ別の細胞に分配され、1本ずつ異なった染色体のセットをもつ生殖細胞が形成される。
この減数分裂の際に性染色体も1本ずつ別の生殖細胞に入る。XYという性染色体をもつ雄ヘテロ型の生物では、雄ではX染色体をもつX精子とY染色体をもつY精子ができる(図1・7)。雌は性染色構成がXXなので、これから形成される卵はすべてX染色体をもつ卵となる。したがって受精の際、X精子がX卵と受精すればXXという雌の子ができるし、Y精子がX卵と受精すればXYという雄の子ができる。すなわち、雄ヘテロ型の生物では、子の性の決定はオスの精子の種類によって、受精の瞬間に決定する。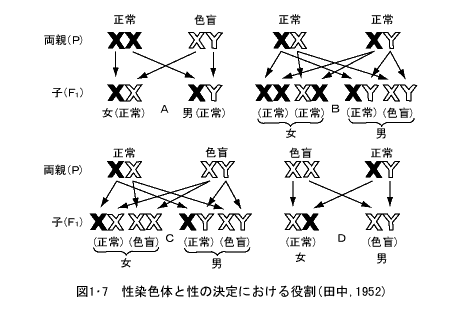
これに対してZWという性染色体をもつ、雌ヘテロ型の生物では、雄ではZZなのでこれから生ずる精子はすべてZ精子のみとなり、卵にZ卵とW卵の2種類ができる。Z卵が受精してできたZZの子は雄、W卵が受精してできたZWの子は雌になり、この場合は卵の種類によって子の性が決定する。
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- 染色体の微細構造
染色体の形と数がそれぞれの生物種によって決まっているのは、すでに述べたとおりである。この染色体は化学的にはデオキシリボ核酸 (DNA)とタンパク質とでできている。 DNAは遺伝子の本体といわれ、後術するように五炭糖(デオキシリボース)とリン酸が交互に連なって鎖状をしたものが2本平行に存在し、その各々の五炭糖に4種類の塩基が一つずつ結合している。さらにこの2本鎖の塩基どうしが互いに結合して2本鎖を結びつけ、全体がらせん状に回転した構造を作っている。
顕微鏡で観察される染色体と、それを構成するDNAやタンパク質との関係はどのようになっているのであろうか。図1・8 に示したように、染色体は動原体の場所でくっついた2本の染色分体(chromatid)からできている。この染色体を酵素などで処理してしだいにほぐしてゆくと、らせん状に巻いたクロマチン繊維となる。このクロマチン繊維からヒストンH1というタンパク質を除去すると、ヌクレオソームというH2A、H2B、H3、H4の4種類のヒストンタンパク質の結合した球状の構造物にDNA分子が巻き付いた基本的な構造となっている。
細胞が分裂するときは核内のDNAがタンパク質と結合して、凝縮して染色体という顕微鏡で観察できる構造物となる。分裂が終わると再び染色体はときほぐれて核内に拡がり、DNA分子までときほぐれてDNAの複製を行う。それが終われば再びタンパク質と結合して凝縮し、染色体としての姿を現す。細胞の分裂、増殖の際にはこのような染色体からDNA分子への拡散、DNA分子から染色体への凝縮というサイクルを繰り返している。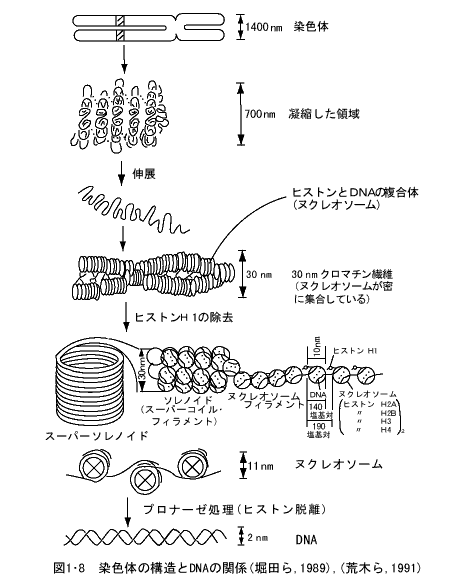
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- 染色体の組換え
生殖細胞ができる時の減数分裂の際に、2本ずつある相同染色体が並び、中間でねじれが起きて、この部分で両染色体が切断し、それぞれ相手の相同染色体の部分と結合して、染色体の組換えが起る。同一染色体上にある二つ以上の遺伝子は、メンデルの独立の法則に従わないで、その染色体と一緒に行動する連関という現象が存在することをすでに述べた(1・4・2)。この連関という現象が染色体の組換えによって破られ、それまで同一染色体上にあって行動を共にしていた二つ以上の遺伝子が、互いに別々の相同染色体に移って、いわゆる遺伝子の組換えが生じる。
一つの染色体上にある二つの遺伝子の間に組換えの起る頻度は、二つの遺伝子間の距離に比例し、両遺伝子間の距離が長いほど、組換えの頻度は高くなり、逆に距離が短いほど遺伝子間の組換え頻度は低い。このような遺伝子間の組換え頻度を調べることにより、遺伝子間の相対的な距離を推定することができる。
さらに一つの染色体上に存在する多くの遺伝子について遺伝子間の相対的な距離の他に、染色体上の遺伝子の位置(遺伝子座という)の順序を知るためには、三点交雑の実験を行う。これは一つの染色体上に存在する三つの遺伝子を使用して交雑実験を行い、その中の二つの遺伝子について、それぞれの相対的な距離を知ることによって、この三つの遺伝子の配列順序を推定することができる(図1・9)。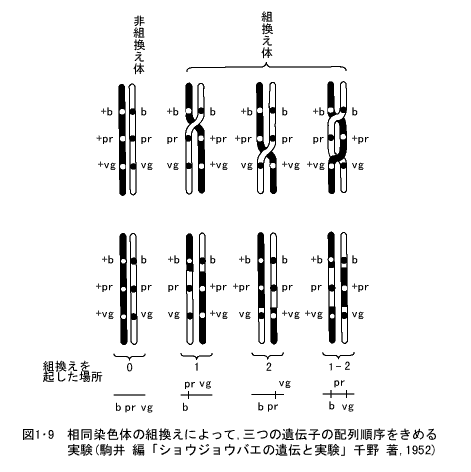
「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
- 染色体地図
ある生物の多くの遺伝的形質について、交配実験を行うことにより、それらの形質を支配する遺伝子の中で、連関現象を示すものがあれば、それらは同一染色体上に存在する遺伝子であることがわかる。また、このようにして交配実験を数多く行うことによって、互いに連関現象を示す遺伝子群(これを連関群という)に区分することができる。 ある生物について、この連関群の数が細胞学的に観察される染色体の半数体(ゲノム)の数と一致する。
このようにして、ある生物について多くの遺伝子の相対的な位置関係を調べ、それを各染色体について示したものが染色体地図(chromosomemap)である。キイロショウジョウバエ(Drosophila melanogaster)では、数千の遺伝子が知られているが、それぞれの遺伝子の染色体上の位置を4対の染色体ごとに示した染色体地図が作成されている(図1・10)。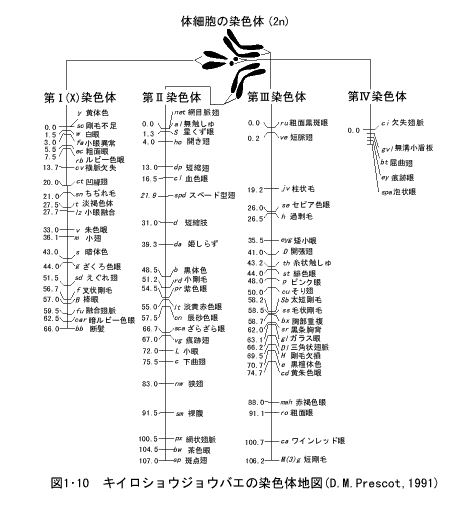
キイロショウジョウバエでは4対の染色体の中で、第1染色体がX染色体(性染色体)で、残りの第2~第4染色体が常染色体である。第1染色体は端部動原体型、第2、第3染色体は中部動原体型であり、第4染色体は点状染色体で遺伝子の数も少ない。
染色体地図の上に記された各遺伝子は複眼や、翅、剛毛の形や色などに関する突然変異の遺伝子を示し、これは自然に生じたものもあるし、X線や化学物質などの処理によって人為的に誘発したものもある。各遺伝子記号はイタリック体の英語で示され、この中で大文字のものは優性、小文字のものは劣性の遺伝子を表す。
キイロショウジョウバエ以外でも、多少方法が異なるものもあるが、ヒトやマウス、ラット、カイコ、線虫などの動物のほか、コメ、ムギ、トウモロコシ、アサガオなどの植物や、酵母やアカパンカビなどの菌類や、大腸菌、サルモネラ菌などの細菌類、バクテリオファージのようなものについても、染色体地図が作成されている。このような染色体地図は新しい突然変異が見つかった場合に、その遺伝子座位を決定するのに役立つほか、これらの遺伝子を用いて、放射線や化学物質による染色体組換えの頻度を調べたり、連関関係を利用して新しい組換え生物を作出したりするのにも有用である。
ヒトの場合には、自由な交配実験ができないために染色体地図の作成が遅れたが、ヒトとマウスの培養細胞を用いて細胞融合を起させ、雑種細胞(hybrid cell)を作成して、この雑種細胞がその後の培養でヒトの染色体だけが脱落して行く現象を利用して、酵素やタンパク質を支配する遺伝子がどの染色体上にあるかが調べられている。また、遺伝子を分離してそのDNAを放射性物質で標識し、ヒトの染色体標本に加えて染色体上の同じ遺伝子DNAと分子雑種を作ることで、遺伝子の座位が決定されている。大腸菌などの細菌の場合には、細菌にも雌雄があることをTatumとLederberg(1946)により発見され、雄の細菌(F+)と雌の細菌(F-)の間に接合が起り、雄の細菌の環状の染色体の1か所が切れて、一方の端から雌の細菌に染色体が移動して行く順序をもとに、染色体地図が作成されている。「基礎遺伝学」(黒田行昭著:近代遺伝学の流れ)裳華房(1995)より転載
