遺伝子の進化とは~ABO式血液型を中心に~
- 遺伝子の系統樹
生物進化の根本は、遺伝子(DNA)が自己複製をして子孫に受け継がれていきながら、突然変異を蓄積して変化してゆくことである。突然変異には、塩基の置換や欠失・挿入など多数の種類がある。図1は、DNAの二重らせん1個から4個のコピーが生じる様子を示している。途中で突然変異が生じて、2個のDNAにそれが引き継がれている。この図をさかさまにすると、樹木が枝葉を茂らせているように見えるので、これを「遺伝子の系統樹」と呼ぶ。系統樹というと、生物種の系統関係を頭に浮かべる人が多いと思うが、実は遺伝子の自己複製を直接反映する遺伝子の系統樹の方が基本なのである。ゲノムのなかのそれぞれの遺伝子が、すべてこのような系統関係の履歴を有しており、それは生命の起源にまで遡るのである。なお遺伝子の進化について興味を持たれた方は、拙著『遺伝子は35億年の夢を見る』(大和書房、1997年)をご覧いただきたい。
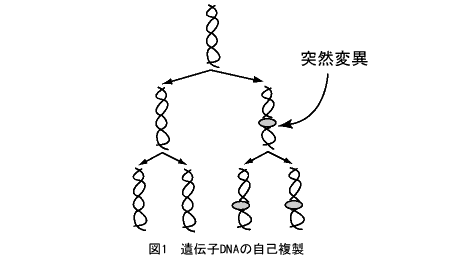
資料:斎藤成也
- ABO式血液型
著者の研究室では、数年前からABO式血液型遺伝子の解析を行なっている。それを以下に紹介しよう。この血液型のA対立遺伝子を持つ人は、からだの中の多数の細胞表面に特有の糖鎖(A型物質)を持ち、B対立遺伝子を持つ人はそれと少し異なるB型物質を持っている。一方、O型の人(O対立遺伝子を2個持つホモ接合型)はA型、B型物質の糖鎖より少し短いH型物質を持っている。つまり、ABO式血液型遺伝子はH型物質の糖鎖に糖をくっつける「糖転移酵素」の遺伝子なのである。
1990年に山本文一郎氏と箱守仙一郎氏らのグループが、A、B、O各対立遺伝子のDNA塩基配列を決定した。その結果、A対立遺伝子から作られるタンパク質とB対立遺伝子から作られるものでは、アミノ酸が4個異なっており、またO対立遺伝子では1塩基の欠失突然変異が生じたためにフレームシフト(遺伝暗号の読み枠がずれること)が起こり、糖転移酵素の働きがなくなっているということがわかった。資料:斎藤成也
- 霊長類のABO遺伝子
ヒト以外の霊長類でもABO式血液型は存在するが、不思議な分布をしている。チンパンジーではA型とO型しかなく、逆にゴリラにはB型だけである。オランウータンや旧世界猿では、A対立遺伝子とB対立遺伝子のどちらもみつかっている。このため、ABO式血液型遺伝子座では、霊長類の進化のかなり古い時代からA・B対立遺伝子が共存してきたのではないかという仮説があった。
この仮説が正しいかどうかは、DNAを調べれば決着するはずである。そこで山本文一郎氏らとドイツの研究グループがあいついで霊長類におけるABO遺伝子座の塩基配列を決定した。図2は、それらの配列データをもとにして作成した遺伝子の系統樹であり、私と山本文一郎氏が共同で発表したものをもとにしてある。この図から、ヒト、ゴリラ、ヒヒで見つかったB型対立遺伝子は、それぞれ独立にA型対立遺伝子から生じたことが予想される(図の点線)。つまり、ヒト、類人猿、旧世界猿の共通祖先では、ABO式血液型の共通祖先遺伝子は、A対立遺伝子タイプであったと考えられる。もっとも、まだ配列はわかっていないものの、オランウータンにもB型が存在するし、ニホンザルも大部分の個体はB型である。このようにあちこちの種でA型とB型が存在していると、祖先型がA型かB型かははっきりしなくなる。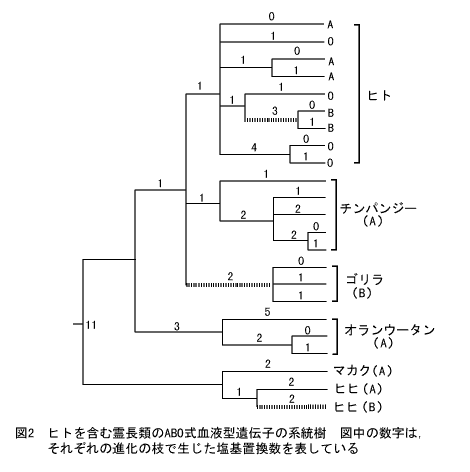
ABO式血液型の遺伝子は、通常の遺伝子がしたがっている進化パターンにはあてはまらないようである。第一に、糖転移酵素の働きがなくなっているのにもかかわらず、O対立遺伝子の頻度がかなり高い。重要な物質交代を担っている酵素遺伝子の場合、酵素の働きがなくなってしまう突然変異が生じると、普通はその個体の生存にきわめて不利なので、子孫を残しにくくなるはずである。したがって、ABO式血液型に関与する糖転移酵素は、人間がとりあえず生きてゆくのには絶対必要であるわけではない。ところが私たちの推定によると、後述するように、ABO式血液型の遺伝子は脊椎動物が出現した5億年以上前ごろから、延々と生き残ってきたのである。このことから、弱いながらもこの遺伝子にはなんらかの存在意義があると考えられる。このように、絶対に必要というわけではないが、あったら少しは役に立つという遺伝子が、ヒトゲノム中にはたくさんあると筆者は考えている。
また、A型とB型の対立遺伝子の共存が霊長類のあちこちの種でみられることも不思議である。この遺伝子がなぜこのような変異パターンを示すのか、まだよくわかっていないが、バクテリアやウイルスなどの感染を防ぐのに、ある程度の効果があるのではないかと考えられている。実際、胃潰瘍や胃癌の原因のひとつであるヘリコバクター=ピロリというバクテリアは、胃壁にもぐりこむ際に、ABO式血液型物質の前駆体であるH型物質を足場にしている。するとH型物質しか持っていないO型の人間は、胃潰瘍などになりやすいため、多少は生存に不利となるだろう。同様のことが、A型やB型の糖抗原と別のバクテリアの組み合わせについても成り立つかもしれない。しかし、まだこれらは仮説にすぎず、将来の検証が待たれるところである。資料:斎藤成也
- ABO遺伝子とGAL遺伝子
突然変異のひとつに「遺伝子重複」があるが、重複したあとの2個の遺伝子コピーがどうなるかを考えてみよう。これらのコピーはそれぞれ独立に塩基置換などの突然変異を蓄積してゆくが、その中には、遺伝子の機能を決定的に損ねてしまうので、普通なら残ることができないものもあるはずである。遺伝子重複が生じる前なら、このような突然変異は著しく生存に不利なので、すぐ消えてしまう。しかし、今やふたつの遺伝子があるので、ひとつにガタがきてももうひとつの遺伝子がしっかりしている限り、そのような突然変異を持った生物個体は大丈夫である。そのため、どちらか片方の遺伝子コピーはその機能を失ってしまうことがあると予想できる。このような遺伝子を、働きを保っている「機能遺伝子」に対して「偽遺伝子」と呼ぶ。偽遺伝子はタンパク質を作り出すことがうまくできないので、その意味では死んでいるが、遺伝子DNAそのものは、ゲノムの中にとどまって親から子へと伝えられてゆく。
偽遺伝子の進化速度が速いことは、中立説から予言されることである(中立説について詳しくは、提唱者の木村資生氏が著した岩波新書「生物進化を考える」を参照されたい)。偽遺伝子がまったく機能を持たないとすれば、そこに生じる突然変異はどのようなものでも生き残ってゆくことができる。まともなタンパク質を作れなくなる強烈な突然変異をくらって死んだ状態になってしまったのだから、そのあとはどのような突然変異を繰り返し受けても、生物個体は平気である。すると、今まで自然淘汰によってすぐ消えていってしまった突然変異も、偽遺伝子では蓄積されることが可能になったので、その分だけ進化速度が大きくなるはずである。実際にそうであることが、現在までに多数の偽遺伝子と機能遺伝子の比較から証明されている。
細胞表面には多数の糖鎖が突き出ている。これらの糖鎖を生み出す糖転移酵素のひとつであるアルファ1・3ガラクトース転位酵素(以下GALと略称)は、脊椎動物が誕生した前後に、遺伝子重複によってABO式血液型遺伝子と分岐しただろうと私たちは推測している。なお、ヒトゲノム中では、どちらの遺伝子も第9番染色体長腕の9q34という番地に位置しており、直列型の遺伝子重複で生じたのは明らかである。
図3はこれら2種類の遺伝子の系統樹である。ヒトゲノムの中にはABO式血液型遺伝子の偽遺伝子が存在することを、山本文一郎氏らが発見したが、この系統樹によると、この偽遺伝子が遺伝子重複を起こして誕生したのは、哺乳類の共通祖先がいた1億年前ごろまで遡るようである。なぜなら、この偽遺伝子が分岐したあとにマウスのABO式血液型遺伝子が枝分かれしているからだ。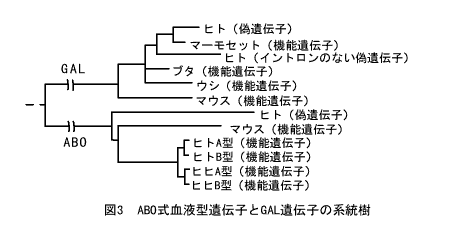
資料:斎藤成也
- GAL遺伝子の偽遺伝子化
GALの遺伝子は、ヒトではだれでも偽遺伝子になっている。こちらの場合はABO式血液型遺伝子の偽遺伝子とは異なり、遺伝子重複が起こることなしに、酵素タンパク質を作っていた本体の機能遺伝子が死んでしまったのだ。一方、マウス、ブタ、ウシなど他の哺乳類はGALの機能遺伝子を持っているのでこの酵素が作られ続けているし、新世界猿であるマーモセットでも機能遺伝子のままである。このほかヒトゲノムには、イントロン部分のないタイプのGAL偽遺伝子も存在する。これはDNAが転写されてメッセンジャーRNAになったあと、逆転写酵素の働きで再びDNAにもどり、それがゲノムの別の場所に挿入されて生じたものである。通常の遺伝子重複で生じる偽遺伝子と違い、いったんメッセンジャーRNAの形になるので、イントロンを失ってしまう。このタイプの遺伝子をプロセス型偽遺伝子と呼ぶ。
霊長類の中でさらにくわしく見てみたのが図4である。この酵素遺伝子は狭鼻猿類(旧世界猿とヒト上科の総称)が広鼻猿類(新世界猿)と枝分かれしたあとに偽遺伝子化していることがわかる。偽遺伝子化したあとは機能を保ったままでいる新世界猿のGAL遺伝子よりも枝が長くなっており、進化速度が速くなっていることがわかる。これは通常の中立進化する偽遺伝子のパターンからの予想と一致している。
ウシやマウスなど他の哺乳類および霊長類でも新世界猿ではGAL遺伝子が生きているのに、人間を含む狭鼻猿の霊長類でGAL遺伝子が死んでしまった(偽遺伝子化した)のはなぜだろう。いくつかの可能性が考えられるが、中立進化を仮定すると、これらの霊長類の系統ではなんらかの理由でGAL遺伝子が働く必要がなくなったということになる。
ところが、偽遺伝子化したあとの進化パターンは中立であっても、細胞表面の糖鎖構造の違いを考えると、次のような仮説が考えられる。狭鼻猿類の祖先が新世界猿の系統と分かれたあと、あるときこの酵素の作り出す糖鎖構造とよく似た糖鎖を持つ悪性のバクテリアに感染されてしまった。大腸菌O157のような毒素を持つバクテリアを考えればよいだろう。このような状況になると、GAL機能遺伝子が死んでしまって偽遺伝子化した個体のほうが、GAL酵素が作り出していた糖鎖に対する抗体を作れるので、このバクテリアを攻撃することができ、より強い免疫力が生じる。GAL酵素を作る個体では、自分と同じ糖鎖抗原を持っているバクテリアは攻撃しにくいからである。この仮説が正しければ、GAL遺伝子が偽遺伝子化した個体のほうが、生存に有利だったことになる。
もちろんこれはひとつの仮説であり、GAL遺伝子はまったく必要がないというわけではないが、あればあったほうがよい、という程度のものなのかもしれない。生物は膨大な進化の歴史をひきずっており、きちんとした設計に基づいて組み立てられているわけではない。したがってこのようなあいまいな機能を持った遺伝子が、ゲノム中から今後多数発見されるだろうと私は考えている。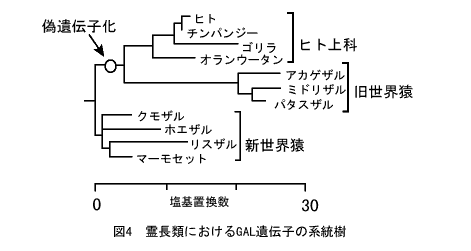
資料:斎藤成也
トピックス
- ABO式血液型遺伝子の系統ネットワーク解析
生物進化の根本は、遺伝子が自己複製をして子孫に受け継がれてゆくあいだに、突然変異を蓄積して変化することである。これによって生じるパターンを「遺伝子の系統樹」と呼ぶ。系統樹というと、生物種の系統関係を頭に浮かべる人が多いと思うが、実は遺伝子の自己複製を直接反映する遺伝子の系統樹のほうが基本なのである。ゲノムの中のそれぞれの遺伝子がすべてこのような系統関係の履歴を有している。現在ではおびただしい数の塩基配列が多数の生物ですでにわかっており、これらを比較することによって遺伝子の系統樹が復元されている。
ヒトのように巨大なゲノムをもつ生物は、長大な染色体を有していることが多いので、染色体の中にずらっと並んでいる遺伝子はひんぱんに組換えや遺伝子変換を行なって新しいハプロタイプ(同じ染色体上に並んでいる遺伝子のつらなり)が生成される。すると、一つ一つの相同な塩基ポジションについては、自己複製の履歴を反映した系統関係が常に存在するが、それらを並べた塩基配列が一つのまとまりとして子孫に伝わるとは必ずしも言えなくなる。この場合には、系統樹の概念を拡張した系統ネットワークを作成してみることが有用である。ABO式血液型遺伝子の進化に系統ネットワークを応用した例を紹介しよう。
ABO式血液型の遺伝子は、糖鎖に糖をくっつける「糖転移酵素」をコードしている。A対立遺伝子をもつ人は、からだの中の多数の細胞表面に特有の糖鎖(A型物質)をもち、B対立遺伝子をもつ人はそれと少し異なるB型物質をもつ。一方、O型の人は酵素活性がないために、A型物質やB型物質の前駆体であるH型物質をもつ。1990年に山本文一郎氏と箱守仙一郎氏らのグループが、ヒトのA、B、O各対立遺伝子のDNA塩基配列を決定した。またヒト以外の霊長類でもABO式血液型遺伝子座の塩基配列がすでにいくつか発表されている。
図(A)は、これら霊長類のデータから作成した遺伝子の系統ネットワークである。全体としては無根系統樹に似ているが、正方形がつながったり立方体が現れているのは系統樹にない特徴である。これは、複数の塩基サイトが互いに矛盾する系統関係を支持する場合に生じる。たとえば、ゴリラ(すべてB型)とヒトのB型には、他の塩基配列と共通に異なっているサイトがあるが、それらは別の塩基サイトでヒトのABO式血液型の遺伝子すべてが共通にもっている塩基の分布パターンと矛盾する。このようなことから、「田」の字のネットワーク構造が生じるのである。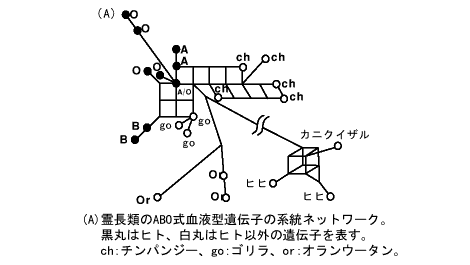
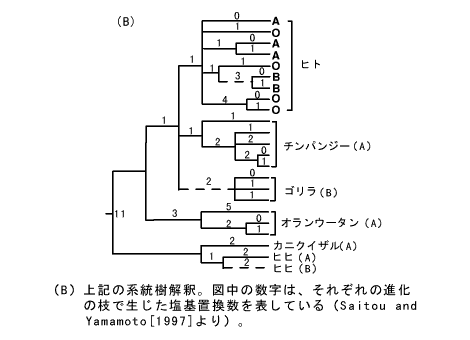
この系統ネットワークには、多数の系統樹が含まれている。その中で、最も確からしいと思われる系統樹を抜き出したものが図(B)である。この系統樹を仮定すると、ヒト、ゴリラ、ヒヒで見つかったB型対立遺伝子は、それぞれ独立にA型対立遺伝子から生じたことが予想される(図の破線)。つまり、ヒト、類人猿、旧世界猿の共通祖先では、ABO式血液型の共通祖先遺伝子は、A対立遺伝子タイプであったと考えられる。もっとも、オランウータンにもB型が存在するし、テナガザルにもA型とB型が存在する。またニホンザルも大部分の個体はB型である。このようにあちこちの種でA型とB型が存在していると、祖先型がA型かB型かはっきりしなくなる。A型とB型の対立遺伝子の共存が霊長類のいろいろな種でみられることは不思議である。この遺伝子がなぜこのような変異パターンを示すのか、まだよくわかっていないが、バクテリアやウイルスなどの感染を防ぐのに、ある程度の効果があるのではないかと考えられている。
資料:斎藤成也
